

各位老师,大家好!我是14号组“铸麦逐梦”的缑金营,根据组长杜立丰老师的安排,今天由我代表小组值日。我们组的成员有张三坤、卫波、秦志英、张中州、望俊森、李继平、徐开杰、李玮、宋瑜龙、韩然、杨子博、何明琦、袁雨豪、崔培涛、温小伟、金燕、杨斌、高宏伟和马宏亮等老师。我们组的杨斌老师上个月分享了“小麦旗叶叶绿素含量遗传分析”一文,介绍了叶绿素含量在小麦产量和育种中的检测与价值。承接杨老师的分享内容,我值日的文章题目是“小麦高光效遗传与种质创新”,继续从小麦光合有关性状的角度讨论小麦产量的调控因素。不妥之处请各位老师批评指正。
我国半个多世纪的小麦生产实践表明,品种遗传改良为我国小麦总产量提升贡献了接近50%。在育种家的不懈努力下,我国小麦产量达到了较高的水平,育种实践中出现了粒数-粒重制约关系:增加一个往往导致另一个因素降低,难以共同提高。小麦单产由单位面积穗数、单穗粒数(前两个因素共同决定粒数)和粒重共同决定。粒重相对稳定,受环境影响小,对我国小麦产量的增加贡献最大,粒重提高是小麦产量改良的重要目标。那么,如何打破粒重-粒数平衡制约关系,同时提高粒重和粒数,进一步提高产量呢?
光合作用可以利用光能固定二氧化碳,将太阳能转化成化学能,为生物圈提供能量,是人类社会赖以生存的基础。任何在作物光合作用方面的改良都有可能导致作物产量的显著提升。因此,提高营养器官光合作用是增加作物产量的理论手段之一,是植物和农业有关基础理论领域研究的热点。作为将太阳光能转化成化学能的主要方式,光合作用的增加有望为储藏器官(如小麦种子)提供更多能量,从而赋予粒重和粒数同时提高的可能,在更高的层次重新平衡。但是,对禾本科农作物的大量研究表明,作物光合效率由多个基因共同调控,优异变异在群体中的分布偏低,利用目前常用的全基因组关联分析定位高光效基因的难度很大。
为了克服上述提高小麦光合作用和产量中遗传材料和关键基因不足的瓶颈,我们计划从人工变异群体出发,寻找与小麦光合和籽粒相关的(CO2 Assimilation Rate and Kernel Enhanced,cake)变异,克隆关键调控基因,然后在天然品种中寻找优异等位变异,创制在光合作用、籽粒甚至产量方面具有优势的种质资源。为此,我们利用化学诱变剂甲基磺酸乙脂(EMS)处理小麦成熟种子,加速自然进化进程,创制人工变异群体。我们课题组获得了春小麦四倍体Kronos和Fielder的突变体库,其中Fielder突变体超过两万个M1单株,预测平均基因组每个GC具有超过一次以上的突变,因此覆盖率很高。我们经过连续八年单行种植和观察,在高度纯合的M8后代群体里面筛选了一批候选突变体。我们对部分个体进行了初步研究,部分结果已经发表,向大家汇报一下。
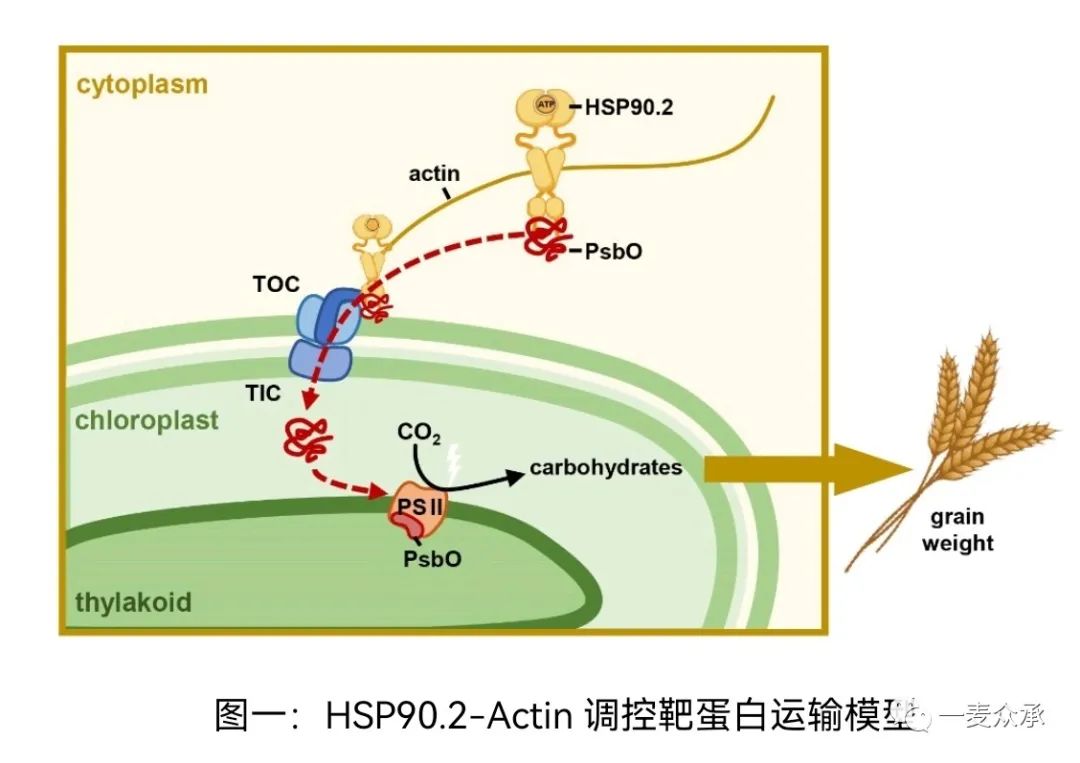
通过对cake1突变体的分析,我们发现热激蛋白(heat shock protein)HSP90.2与光合效率和粒重正相关,HSP90.2功能完全丧失可以导致产量下降80%左右,相反过量表达增强HSP90.2功能可以显著提高光合作用、粒重和产量。作为真核细胞中普遍存在的具有高度保守的分子伴侣,HSP90.2定位在细胞质,结合一系列细胞核编码的光系统蛋白,如外周蛋白PsbO。hsp90.2突变体或者RNAi转基因小麦的叶绿体中,PsbO积累减少,而HSP90抑制剂处理也可以减少目的蛋白的积累,暗示HSP90.2对靶蛋白的定位至关重要。前人研究表明,90%以上的叶绿体蛋白在细胞核中编码,翻译后转运进入叶绿体。因此,利用HSP90.2增强光系统成员靶向运输是提高植物产量的关键(图一)。
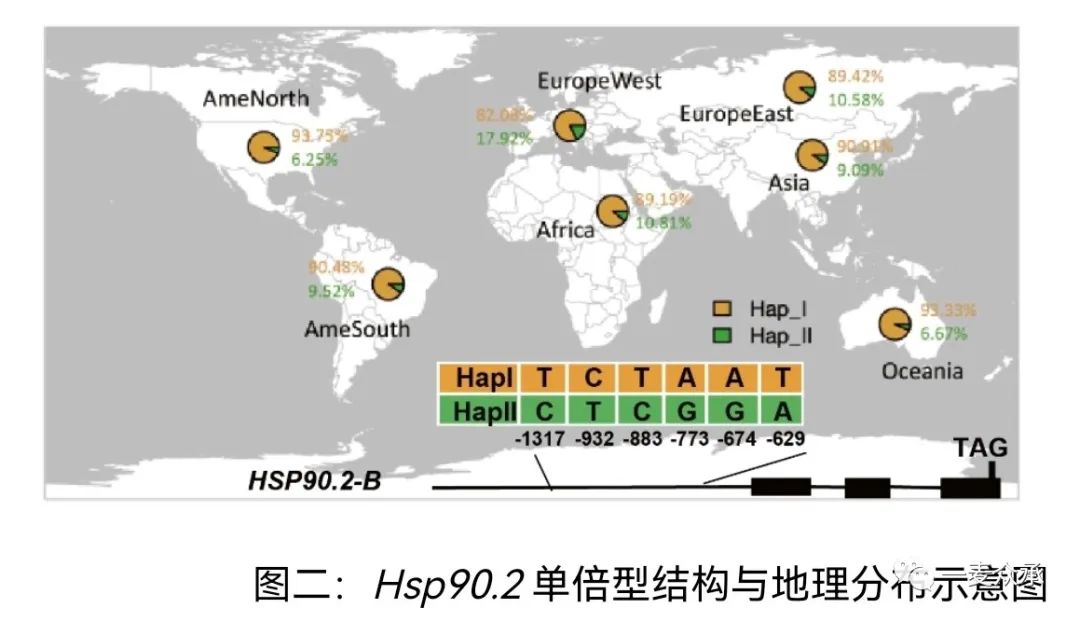
为了研究HSP90.2在小麦生产中的应用价值,我们与河南农业大学陈锋教授合作,分析了自然群体中该基因的多态性。根据小麦重测序数据,HSP90.2在启动子上具有多个单倍型,其中单倍型II(Hsp90.2-HapII)携带六个单核苷酸多态性(SNP)位点,在RIL群体中,Hsp90.2-HapII显著提高HSP90.2的基因表达和蛋白含量,增强了光合效率,促进了产量的提升。
值得指出的是,由于以往基础研究不足,Hsp90.2-HapII在育种并未充分被充分选择。在世界范围内,约10%的育成品种携带Hsp90.2-HapII的单倍型,在西欧等发达国家这一比例可以达到18%,但是在巴基斯坦这一小麦单产相对较低的国家,仅有2%的品种携带Hsp90.2-HapII优异单倍型(图二)。我们将两种单倍型的代表性品种杂交,获得了双亲分离群体,发现携带Hsp90.2-HapII优异单倍型的个体在光合效率、粒重和产量方面具有明显的优势。相关论文今年初发表在《Plant Biotechnology Joural》杂志,请大家批评指正。此外,Hsp90.2-HapII优异单倍型同时可以提高小麦抗病能力,请参考我们近期发表在《Plant Cell & Environments》杂志的论文。通过陈红敏老师组织的“一麦众承”种质资源分享活动,我们分享了携带Hsp90.2-HapII优异单倍型的材料,希望各位老师在育种实践中使用,分析这个单倍型是否具有育种价值,以及适合的环境。
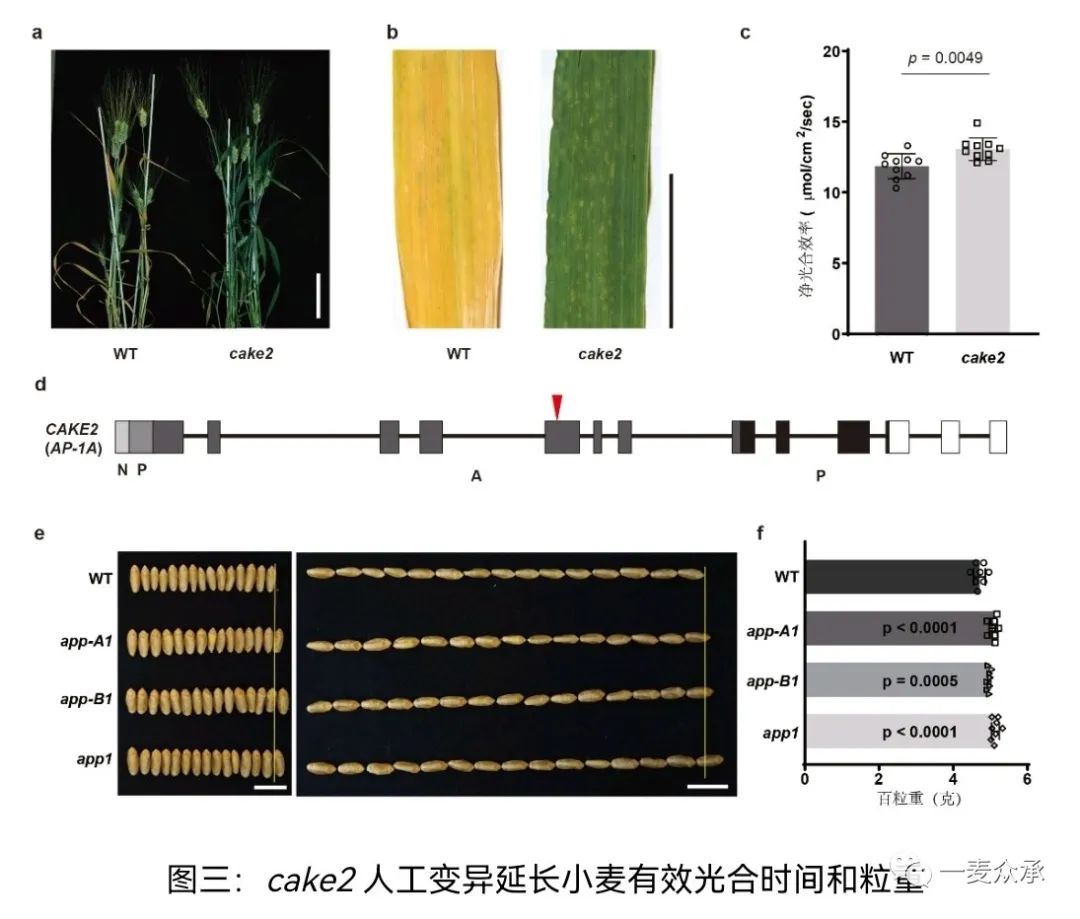
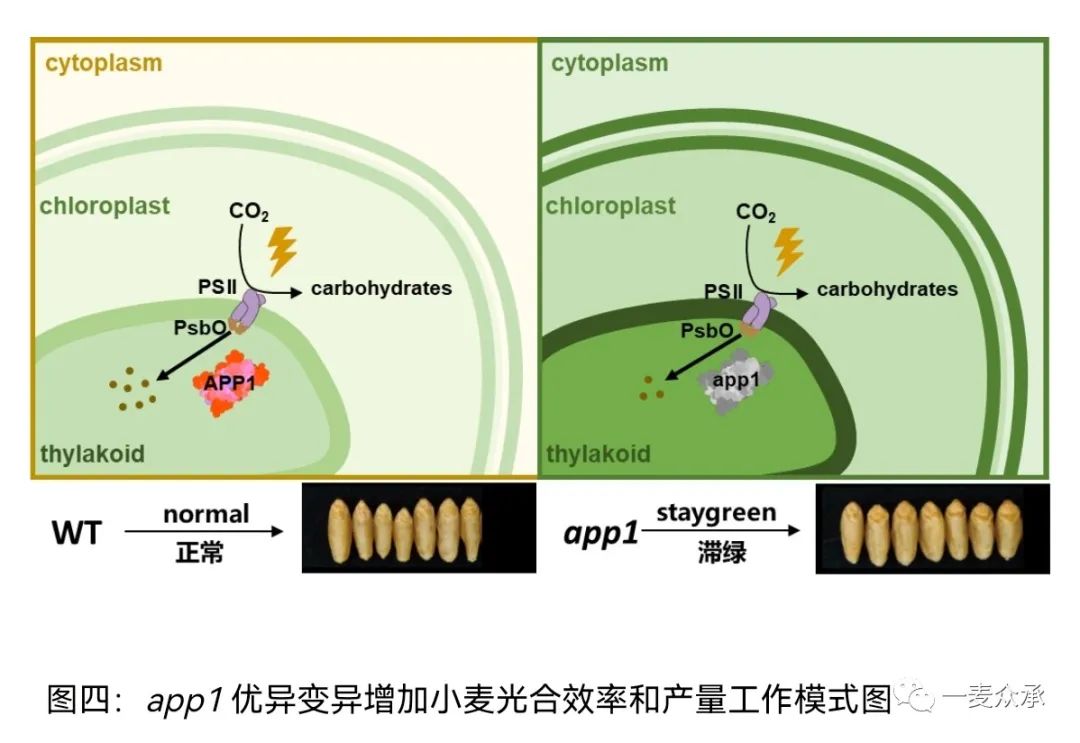
cake2突变体有效光合持续时间延长,籽粒变大,粒重增加,产量提升显著。遗传学分析表明:cake2的A基因组中天冬氨酸蛋白酶基因(Asppartic Protease1, APP1)编码区具有一个提前终止的突变,编码的蛋白丢失了一个酶活重要结构域。随后研究表明,小麦B基因组的功能抑制突变也可以增加光合效率和产量,双突变体在光合效率和粒重方面具有明显的优势(图三)。在植物体内,APP1蛋白定位于叶绿体,正在酵母双杂交系统中可以结合光系统II的外周蛋白PsbO,后者可以保护光系统II。在app1突变体中PsbO的稳定性显著提高,完整的光系统蛋白复合体比例更高,与功能性滞绿的现象一致。
为了寻找在育种过程中小麦种质资源APP1可以应用育种实践的自然变异,我们分析了小麦重测序数据,发现APP1蛋白第442位为氨基酸一般是丝氨酸(单倍型I,HapI),但是少数品种为甘氨酸(单倍型II,HapII),会导致APP1蛋白丰度降低。我们广泛收集了携带单倍型II的品种,与单倍型I的品种比较粒重增加,产量具有优势。为了进一步研究APP1单倍型在其中的贡献,我们将代表性品种扬麦5号(单倍型I)和郑麦9023(单倍型II)杂交,创制了双亲分离群体,在田间种植中,携带APP1单倍型II的后代在光合效率和粒重等方面具有明显的优势,表明APP1单倍型II可以作为一个分子标记由于育种实践,提高小麦单产。相关论文今年六月份发表在《Nature Plants》杂志,请大家批评指正。
除了上述已经发表的工作,我们也克隆了其他几个基因:cake3可以通过增加光合源提高小麦粒重和产量,增强小麦抗条锈病的能力,使籽粒蛋白增加1%左右;cake4可以改良株型,提高冠层截获光的效率,增加产量,提高籽粒抗氧化活性,耐盐能力显著增强;CAKE5调控碳代谢途径,与CAKE1形成蛋白复合体,增强CAKE1的功能,提高小麦粒重和抗病能力。大家如果感兴趣,我们可以讨论并开展合作,将基础研究和育种实践结合,推动小麦产业的整体发展,利国利民,体现我们科研活动的价值。
作者简介:缑金营,男,博士,教授,博导,在中国农业大学农学院小麦研究中心工作,手机15921835089,邮箱jygou@cau.edu.cn。主要从事小麦高产高光效及抗条锈病分子遗传研究。在《Nature Plants》、《Nature Communications》和《Plant Cell》等期刊以第一或通讯作者发表论文30 余篇,总引用2300 多次,单篇引用最高超过700 次。目前担任《BMC Plant Biology 》和《Molecular Breeding 》杂志编委。
